

Lee, Comparte y únete a la pagina y grupo de Facebook…
Pagina en Facebook
https://www.facebook.com/La-cosa-aquella-231455250559123/
Grupo de Facebook
https://www.facebook.com/groups/1013342072090602/
Blog
https://kenzocaspi.wordpress.com/author/kenzocaspi/
Canal Youtube
https://www.youtube.com/channel/UCJZIxTmbZ4CwPCE0Y4UAI1g
Por Gordon Duff, editor sénior -17 de mayo de 2020
Este artículo contiene pruebas que no pueden ser cuestionadas o denegadas, que puede enviar a cualquier agencia gubernamental o profesional de la salud.
Lo que aún no está probado, pero se está enfocando en que el programa de armas biológicas de EE. UU. En Fort Detrick, Maryland, el equipo y ciertamente el personal clave, ciertamente migraron a laboratorios secretos en las grandes universidades estatales para «esconderse a simple vista».
Siga las carreras, todos los enlaces están incluidos, de aquellos que trabajaron en el proyecto Wuhan-COVID en 2017.
Además, tenga en cuenta que se utiliza exactamente el mismo personal y equipo para la investigación y las pruebas de «prevención» falsas que para la fabricación de armas y la producción real.
Desde que se escribió este artículo, hemos comenzado a analizar las operaciones mundiales del contratista nuclear / bio / químico estadounidense, el favorito de Kushner-Trump, Battelle, y sus laboratorios secretos en todo el mundo.
Cuando comenzamos, nuestra gente comenzó a ser amenazada. Ese fue un grave error.
Envíe este documento a cualquier médico u otro especialista calificado en biociencias. Mira lo que dicen.
Introducción
Los documentos a continuación mostrarán que la investigación para crear COVID 19 comenzó en los Estados Unidos en 2006 y culminó en un exitoso arma biológica en 2015, con un trabajo realizado en la Universidad de Carolina del Norte y en Harvard y en el laboratorio de la Administración de Alimentos y Medicamentos de Arkansas .
Su trabajo se tituló:
Un grupo de coronavirus de murciélago circulante similar al SARS muestra potencial para la emergencia humana
Hicieron esto y más, tanto más como leerán a continuación.
Como dijo Trump, una y otra y otra vez, los chinos estaban involucrados.
El Laboratorio clave de patógenos especiales y bioseguridad, el Instituto de Virología de Wuhan, la Academia de Ciencias de China, Wuhan, China, suministró el virus del murciélago de Wuhan que se utilizó en el estudio estadounidense. Su nombre fue incluido solo por esa razón.
COVID 19 fue un proyecto de armas biológicas del ejército de los EE. UU. Para fabricar una enfermedad que causa neumonía y que sería casi imposible vacunar en pacientes mayores de 40 años.
La prueba está aquí. El estudio fue dirigido por la Universidad de Carolina del Norte y financiado por USAID / CIA. Eligió un virus de murciélago chino y también eligió incluir un centro médico en Wuhan.
Ahora sabemos por qué, una cortina de humo de la culpa de un programa con el que China tenía poco o nada que ver, algo satánicamente malvado y puramente estadounidense.
En noviembre de 2015, se publicó un estudio que describe la capacidad de producir el virus que estamos tratando ahora. Entre los muchos involucrados había un laboratorio en Wuhan, China. Fue catalogado desde el principio como una de las docenas, en su mayoría estadounidenses, que trabajan en este proyecto.
Sin embargo, un participante clave quedó fuera, USAID. Se sospecha, profundamente, que USAID es un frente para la investigación estadounidense de bioguerra como la realizada en Tbilisi, Georgia y en otros lugares, muy documentada. Esta es la cita que agrega USAID al grupo de financiación de la investigación.
Cambia la historia
20 de noviembre de 2015
En la versión de este artículo publicada inicialmente en línea, los autores omitieron reconocer una fuente de financiamiento, financiamiento de USAID-EPT-PREDICT de EcoHealth Alliance, a Z.-LS. El error se ha corregido para las versiones impresas, PDF y HTML de este artículo. .
Ahora presentaremos el artículo sesgado de Pravda y, debajo de eso, el estudio real que demuestra la capacidad de producir COVID 19, que demuestra que no es un virus natural de una vez por todas.
En cuanto a quién hizo qué, este no es nuestro trabajo, pero estamos demostrando, categóricamente, que cuando se menciona un laboratorio chino, es un jugador menor en un esfuerzo estadounidense, como se describe exhaustivamente a continuación.
Esto hace que discutir el laboratorio de Wuhan sea posiblemente cómplice de la bioguerra.
De manera similar, cuando la revista Forbes y otros declararon que podían probar que COVID 19 se hizo de forma natural y, por supuesto, tenían el mismo acceso que nosotros, sospechamos que son parte de un esfuerzo de desinformación vinculado a USAID y la guerra biológica.
La sospecha no es prueba. La prueba es prueba y hay pruebas suficientes para ahogarse. Nuestro agradecimiento a los profesionales médicos estadounidenses que se lanzaron al Ejército de EE. UU. Y a la CIA y que ayudaron a llevarnos a donde estamos ahora, una nación hecha pedazos.
Pravda.Ru: dicho material apareció en 2015 en el sitio web de la revista científica Natura en 2015. Luego, los autores afirmaron que después de la aparición del virus del SARS (2002-2003) y el síndrome respiratorio del Medio Oriente (MERS), los científicos sabían del riesgo de transmisión interespecífica que conduciría a una epidemia entre las personas.
Experimento exitoso de laboratorio
Entre otras cosas, el equipo de investigación estudió los murciélagos, que son las incubadoras más grandes de coronavirus. Sin embargo, los murciélagos no podían transmitir el coronavirus a los humanos porque no podían interactuar con las células humanas con receptores ACE2.
El material también indicó que los murciélagos en herradura llevan una cepa de coronavirus SARS que puede transmitirse a los humanos. Se le ha denominado el virus SHC014-CoV.
Para estudiar mejor este virus, los científicos copiaron el coronavirus y lo infectaron con ratones de laboratorio. Los resultados mostraron que el virus es realmente capaz de unirse a las células humanas con receptores ACE2 y multiplicarse en las células del sistema respiratorio.
En el trabajo de investigación, se observa que los materiales de laboratorio, las muestras y el equipo que se utilizaron en la investigación se obtuvieron del Instituto de Investigación Médica del Ejército de Enfermedades Infecciosas. Aunque todavía no es posible decir con certeza que el virus que se probó en ratones de laboratorio es el mismo que el coronavirus SARS-Cove-2.
Política de la OTAN
Sin embargo, se pueden encontrar cosas interesantes en documentos anteriores.
Por ejemplo:
- El informe de actividades de la Alianza de 2019 dice que en 2019, el primer lugar de la Alianza en investigación y desarrollo estuvo ocupado por el tema de la protección radioquímica y biológica (29%), cambiando el problema aparentemente más acuciante de Europa: el contraterrorismo (resultó ser 4 – m prioridad).
- Un año antes, en 2018, la situación era exactamente la opuesta: el terrorismo, como debería ser, estaba en primer lugar (28%) y la protección radioquímica y biológica en el cuarto (13%).
Como escribe la snitch de Bruselas en el canal de telegramas, «dada la ausencia de razones visibles para un cambio tan brusco en los intereses científicos, hay dos opciones y ambas son desagradables:
- o la OTAN ahora muestra el quinto punto, falsificando los datos para mostrar «y siempre estamos preparados para virus, somos modernos»,
- o incluso en 2019 en la alianza, Dios me perdone, sabían de dónde vendrían los problemas.
Sí, la primera opción es mucho más real, pero, como ve, los hechos son sorprendentes.
Fuente: Pravda.
Investigación original de 2015 sin editar y completa
Nature Medicine, volumen 21 , páginas 1508–1513 ( 2015 )
El 6 de abril de 2016 se publicó una corrección de errores de este artículo.
Resumen
La aparición del coronavirus del síndrome respiratorio agudo severo (SARS-CoV) y el síndrome respiratorio del Medio Oriente (MERS) -CoV subraya la amenaza de eventos de transmisión entre especies que provocan brotes en humanos. Aquí examinamos el potencial de enfermedad de un virus similar al SARS, SHC014-CoV, que actualmente circula en las poblaciones de murciélagos de herradura chinos 1 . Usando el sistema de genética inversa SARS-CoV 2 , generamos y caracterizamos un virus quimérico que expresa el pico del coronavirus de murciélago SHC014 en un esqueleto de SARS-CoV adaptado a ratón.
Los resultados indican que los virus del grupo 2b que codifican el pico SHC014 en una columna vertebral de tipo salvaje pueden usar de manera eficiente múltiples ortólogos de la enzima convertidora de angiotensina humana del receptor del SARS II (ACE2), replicarse eficientemente en células primarias de las vías respiratorias humanas y lograr títulos in vitro equivalentes a cepas epidémicas de SARS-CoV. Además, los experimentos in vivo demuestran la replicación del virus quimérico en pulmón de ratón con notable patogénesis.
La evaluación de las modalidades inmunoterapéuticas y profilácticas basadas en el SARS reveló una eficacia deficiente; tanto el enfoque de anticuerpos monoclonales como el de vacunas no lograron neutralizar y proteger contra la infección con CoV utilizando la nueva proteína espiga.
Sobre la base de estos hallazgos, hemos derivado sintéticamente un virus recombinante SHC014 infeccioso de longitud completa y demostramos una replicación viral robusta tanto in vitro como in vivo . Nuestro trabajo sugiere un riesgo potencial de reaparición del SARS-CoV de virus que circulan actualmente en poblaciones de murciélagos.
Principal
La aparición del SARS-CoV anunció una nueva era en la transmisión entre especies de enfermedades respiratorias graves con la globalización que conduce a una rápida propagación en todo el mundo y un impacto económico masivo 3 , 4 . Desde entonces, varias cepas, incluidas las cepas de influenza A H5N1, H1N1 y H7N9, y MERS-CoV, han surgido de poblaciones animales, causando enfermedades, mortalidad y dificultades económicas considerables para las regiones afectadas 5 . Aunque las medidas de salud pública pudieron detener el brote de SARS-CoV 4 , estudios recientes de metagenómica han identificado secuencias de virus similares al SARS que están estrechamente relacionados y que circulan en las poblaciones de murciélagos chinos que pueden representar una amenaza futura 1 , 6 .
Sin embargo, la secuencia de datos por sí sola proporciona información mínima para identificar y prepararse para futuros virus pre-pandémicos. Por lo tanto, para examinar el potencial de emergencia (es decir, el potencial de infectar a los humanos) de los CoV de murciélago circulantes, creamos un virus quimérico que codifica una nueva proteína de pico de CoV zoonótica, de la secuencia RsSHC014-CoV que se aisló de los murciélagos de herradura chinos 1 —En el contexto de la red troncal adaptada al ratón SARS-CoV. El virus híbrido nos permitió evaluar la capacidad de la nueva proteína espiga para causar enfermedad independientemente de otras mutaciones adaptativas necesarias en su columna vertebral natural.
Usando este enfoque, caracterizamos la infección por CoV mediada por la proteína de la espiga SHC014 en las células de las vías respiratorias humanas primarias e in vivo y probamos la eficacia de las terapias inmunes disponibles contra SHC014-CoV. En conjunto, la estrategia traduce los datos de metagenómica para ayudar a predecir y prepararse para futuros virus emergentes.
Las secuencias de SHC014 y el RsWIV1-CoV relacionado muestran que estos CoV son los parientes más cercanos a las cepas epidémicas de SARS-CoV ( Fig. 1a, b ); sin embargo, existen diferencias importantes en los 14 residuos que se unen al ACE2 humano, el receptor del SARS-CoV, incluidos los cinco que son críticos para el rango de hospedantes: Y442, L472, N479, T487 e Y491 (ref. 7 ).
En WIV1, tres de estos residuos puede variar en la epidemia cepa Urbani SARS-CoV, pero no se espera que altere la unión a ACE2 ( complementario Fig. 1a, b y la Tabla Suplementaria 1 ). Este hecho es confirmado por ambos experimentos pseudotipificación que midieron la capacidad de los lentivirus que codifican proteínas WIV1 de punta para entrar en las células que expresan humano ACE2 ( complementario Fig. 1 ) y por in vitro ensayos de replicación de WIV1-CoV (Ref. 1 ). Por el contrario, 7 de los 14 residuos de interacción ACE2 en SHC014 son diferentes de los del SARS-CoV, incluidos los cinco residuos críticos para el rango del huésped ( Figura 1 y Tabla 1 suplementaria ).
Estos cambios, junto con la falla de los lentivirus pseudotipados que expresan el pico SHC014 para ingresar a las células ( Figura complementaria 1d ), sugirieron que el pico SHC014 no puede unirse a ACE2 humano. Sin embargo, se informó que cambios similares en las cepas relacionadas con el SARS-CoV permitieron la unión de ACE2 7 , 8 , lo que sugiere que se requieren pruebas funcionales adicionales para la verificación.
Por lo tanto, sintetizamos el pico SHC014 en el contexto de la estructura principal SARS-CoV adaptada a ratón y competente para la replicación (en adelante, nos referiremos al CoV quimérico como SHC014-MA15) para maximizar la oportunidad de estudios de patogénesis y vacuna en ratones ( Figura complementaria . 2a ). A pesar de las predicciones de los experimentos de modelado y pseudotipado basados en estructuras, SHC014-MA15 fue viable y se repitió a títulos altos en células Vero ( Figura complementaria 2b ). Similar al SARS, SHC014-MA15 también requería una molécula funcional de ACE2 para la entrada y podía usar ortólogos de ACE2 humanos, de civeta y de murciélago ( Figura complementaria 2c, d ).
Para probar la capacidad de la espiga SHC014 para mediar la infección de la vía aérea humana, examinamos la sensibilidad de la línea celular epitelial humana Calu-3 2B4 (ref. 9 ) a la infección y encontramos una replicación robusta de SHC014-MA15, comparable a la de SARS-CoV Urbani ( Fig. 1c ). Para extender estos hallazgos, los cultivos primarios de epitelio de las vías respiratorias humanas (HAE) se infectaron y mostraron una replicación robusta de ambos virus ( Fig. 1d ). Juntos, los datos confirman la capacidad de los virus con el pico SHC014 para infectar las células de las vías respiratorias humanas y subrayan la amenaza potencial de transmisión entre especies de SHC014-CoV.
( a ) Las secuencias del genoma de longitud completa de CoV representativos se alinearon y se mapearon filogenéticamente como se describe en los Métodos en línea . La barra de escala representa las sustituciones de nucleótidos, con solo un soporte de arranque superior al 70% marcado. El árbol muestra los CoV divididos en tres grupos filogenéticos distintos, definidos como α-CoV, β-CoV y γ-CoV. Los grupos de subgrupos clásicos se marcan como 2a, 2b, 2c y 2d para los β-CoV y como 1a y 1b para los α-CoV. ( b ) Las secuencias de aminoácidos de los dominios S1 de los picos de β-CoV representativos del grupo 2b, incluido el SARS-CoV, se alinearon y se mapearon filogenéticamente. La barra de escala representa las sustituciones de aminoácidos. ( c , d) Replicación viral de SARS-CoV Urbani (negro) y SHC014-MA15 (verde) después de la infección de células Calu-3 2B4 ( c ) o cultivos de células HAE de interfaz aire-líquido primario bien diferenciados ( d ) en una multiplicidad de infección (MOI) de 0.01 para ambos tipos de células. Se recogieron muestras en puntos de tiempo individuales con repeticiones biológicas ( n = 3) para los experimentos de Calu-3 y HAE. ( e , f ) Pérdida de peso ( n = 9 para SARS-CoV MA15; n = 16 para SHC014-MA15) ( e ) y replicación viral en los pulmones ( n = 3 para SARS-CoV MA15; n = 4 para SHC014- MA15) ( f) de ratones BALB / c de 10 semanas de edad infectados con 1 × 10 4 pfu de SARS-CoV MA15 (negro) o SHC014-MA15 (verde) adaptado a ratón a través de la ruta intranasal (in). ( g, h ) Se muestran imágenes representativas de secciones de pulmón teñidas para el antígeno SARS-CoV N de ratones infectados con SARS-CoV MA15 ( n = 3 ratones) ( g ) o SHC014-MA15 ( n = 4 ratones) ( h ). Para cada gráfico, el valor central representa la media del grupo, y las barras de error definen las barras de Escala sem, 1 mm.
Para evaluar el papel del pico SHC014 en la mediación de la infección in vivo , infectamos ratones BALB / c de 10 semanas de edad con 10 4 unidades formadoras de placa (pfu) de SARS-MA15 o SHC014-MA15 ( Fig. 1e-h ) Los animales infectados con SARS-MA15 experimentaron una rápida pérdida de peso y letalidad 4 días después de la infección (ppp); en contraste, la infección por SHC014-MA15 produjo una pérdida de peso sustancial (10%) pero ninguna letalidad en ratones ( Fig. 1e ). El examen de la replicación viral reveló títulos virales casi equivalentes de los pulmones de ratones infectados con SARS-MA15 o SHC014-MA15 ( Fig. 1f ). Mientras que los pulmones de los ratones infectados con SARS-MA15 mostraron una tinción robusta tanto en los bronquiolos terminales como en el parénquima pulmonar 2 ppp ( Fig. 1g), los de ratones infectados con SHC014-MA15 mostraron una tinción reducida del antígeno de la vía aérea ( Fig. 1h ); en contraste, no se observó ningún déficit en la tinción de antígeno en el parénquima o en la puntuación general de la histología, lo que sugiere una infección diferencial del tejido pulmonar para SHC014-MA15 ( Tabla complementaria 2 ). Luego analizamos la infección en animales más susceptibles, de edad avanzada (12 meses). Los animales infectados con SARS-MA15 perdieron peso rápidamente y sucumbieron a la infección ( Figuras suplementarias 3a, b ). La pérdida de peso robusta y sostenida inducida por la infección por SHC014-MA15, pero tenía una letalidad mínima. Las tendencias en la histología y los patrones de tinción de antígeno que observamos en ratones jóvenes se conservaron en los animales más viejos ( Tabla complementaria 3) Se excluyeron la posibilidad de que SHC014-MA15 se mediar la infección a través de un receptor alternativo sobre la base de experimentos usando Ace2 – / – ratones, que no mostró pérdida de peso o la tinción de antígeno después de la infección SHC014-MA15 ( complementario figura 4a, b. Y complementaria Tabla 2 ). Juntos, los datos indican que los virus con el pico SHC014 son capaces de inducir la pérdida de peso en ratones en el contexto de un esqueleto virulento de CoV.
Dada la eficacia preclínica de las terapias con anticuerpos monoclonales contra el Ébola, como ZMApp 10 , luego buscamos determinar la eficacia de los anticuerpos monoclonales contra el SARS-CoV contra la infección por SHC014-MA15. Anteriormente se habían informado cuatro anticuerpos monoclonales humanos ampliamente neutralizantes dirigidos contra la proteína del pico del SARS-CoV y son reactivos probables para la inmunoterapia 11 , 12 , 13 . Examinamos el efecto de estos anticuerpos en la replicación viral (expresado como porcentaje de inhibición de la replicación viral) y descubrimos que, mientras que el SARS-CoV Urbani de tipo salvaje estaba fuertemente neutralizado por los cuatro anticuerpos a concentraciones de anticuerpos relativamente bajas ( Fig. 2a-d), la neutralización varió para SHC014-MA15. Fm6, un anticuerpo generado por la presentación en fago y los mutantes de escape 11 , 12 , logró solo niveles de inhibición de fondo de la replicación SHC014-MA15 ( Fig. 2a ). De manera similar, los anticuerpos 230.15 y 227.14, que se derivaron de las células B de memoria de pacientes infectados con SARS-CoV 13 , tampoco pudieron bloquear la replicación de SHC014-MA15 ( Fig. 2b, c) Para los tres anticuerpos, las diferencias entre las secuencias de aminoácidos de pico de SARS y SHC014 correspondieron a cambios de residuos directos o adyacentes encontrados en mutantes de escape de SARS-CoV (fm6 N479R; 230.15 L443V; 227.14 K390Q / E), lo que probablemente explica la ausencia de los anticuerpos ‘actividad neutralizante contra SHC014. Finalmente, el anticuerpo monoclonal 109.8 pudo lograr una neutralización del 50% de SHC014-MA15, pero solo a altas concentraciones (10 μg / ml) ( Fig. 2d ). Juntos, los resultados demuestran que los anticuerpos ampliamente neutralizantes contra el SARS-CoV solo pueden tener una eficacia marginal contra las cepas emergentes de CoV similares al SARS, como SHC014.
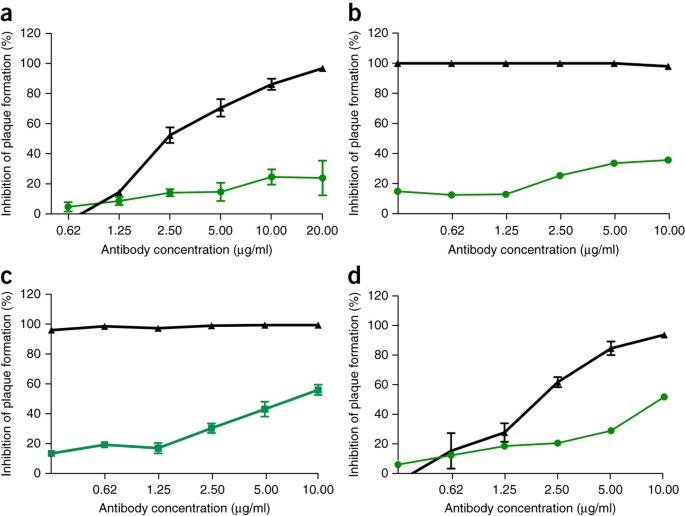
( a – d ) Ensayos de neutralización que evalúan la eficacia (medida como una reducción en el número de placas) de un panel de anticuerpos monoclonales, que originalmente se generaron contra la epidemia de SARS-CoV, contra la infección de las células Vero con SARS-CoV Urbani (negro ) o SHC014-MA15 (verde). Los anticuerpos probados fueron fm6 ( n = 3 para Urbani; n = 5 para SHC014-MA15) 11 , 12 ( a ), 230.15 ( n = 3 para Urbani; n = 2 para SHC014-MA15) ( b ), 227.15 ( n = 3 para Urbani; n = 5 para SHC014-MA15) ( c ) y 109.8 ( n = 3 para Urbani; n = 2 para SHC014-MA15) 13 ( d ). Cada punto de datos representa la media del grupo y las barras de error definen la sem. Tenga en cuenta que las barras de error en las células Vero infectadas con SARS-CoV Urbani en b , c están superpuestas por los símbolos y no son visibles.
Para evaluar la eficacia de las vacunas existentes contra la infección con SHC014-MA15, vacunamos ratones viejos con SARS-CoV (DIV) completo inactivado doblemente. El trabajo previo demostró que DIV podía neutralizar y proteger a los ratones jóvenes del desafío con un virus homólogo 14 ; sin embargo, la vacuna no logró proteger a los animales de edad avanzada en los que también se observó patología inmune aumentada, lo que indica la posibilidad de que los animales resulten dañados debido a la vacunación 15 . Aquí encontramos que DIV no proporciona protección contra el desafío con SHC014-MA15 con respecto a la pérdida de peso o el título viral ( Suplemento Fig. 5a, b ). De acuerdo con un informe anterior con otros grupos heterólogos 2b CoVs 15, el suero de ratones envejecidos vacunados con DIV tampoco logró neutralizar SHC014-MA15 ( Figura complementaria 5c ). En particular, la vacuna DIV resultó en una patología inmune robusta ( Tabla complementaria 4 ) y eosinofilia ( Figura complementaria 5d-f ). Juntos, estos resultados confirman que la vacuna DIV no sería protectora contra la infección con SHC014 y posiblemente podría aumentar la enfermedad en el grupo de ancianos vacunados.
A diferencia de la vacunación de ratones con DIV, el uso de SHC014-MA15 como vacuna viva atenuada mostró una posible protección cruzada contra el desafío con SARS-CoV, pero los resultados tienen advertencias importantes. Infectamos ratones jóvenes con 10 4 pfu de SHC014-MA15 y los observamos durante 28 días. Luego desafiamos a los ratones con SARS-MA15 en el día 29 ( Figura complementaria 6a ). La infección previa de los ratones con la dosis alta de SHC014-MA15 confirió protección contra el desafío con una dosis letal de SARS-MA15, aunque solo hubo una respuesta mínima de neutralización de SARS-CoV del antisuero provocada 28 días después de la infección por SHC014-MA15 ( Suplementario Fig. 6b, 1: 200). En ausencia de un refuerzo secundario de antígeno, 28 ppp representa el pico esperado de títulos de anticuerpos e implica que habrá una protección disminuida contra el SARS-CoV con el tiempo 16 , 17 . Se observaron resultados similares que muestran protección contra el desafío con una dosis letal de SARS-CoV en ratones BALB / c de edad avanzada con respecto a la pérdida de peso y la replicación viral ( Figura complementaria 6c, d ). Sin embargo, la dosis de infección por SHC014-MA15 de 10 4 ufp indujo una pérdida de peso> 10% y letalidad en algunos animales de edad avanzada ( Fig. 1 y Fig. Suplementaria 3).) Descubrimos que la vacunación con una dosis más baja de SHC014-MA15 (100 pfu) no indujo la pérdida de peso, pero tampoco protegió a los animales de edad avanzada de un desafío de dosis letal de SARS-MA15 ( Figura complementaria 6e, f ). Juntos, los datos sugieren que el desafío SHC014-MA15 puede conferir protección cruzada contra el SARS-CoV a través de epítopos conservados, pero la dosis requerida induce patogénesis e impide su uso como vacuna atenuada.
Una vez establecido que el pico SHC014 tiene la capacidad de mediar la infección de células humanas y causar enfermedad en ratones, a continuación sintetizamos un clon infeccioso SHC014-CoV de longitud completa basado en el enfoque utilizado para el SARS-CoV ( Fig.3a ) 2 . La replicación en células Vero no reveló ningún déficit para SHC014-CoV en relación con el del SARS-CoV ( Fig. 3b ); sin embargo, SHC014-CoV se atenuó significativamente ( P <0.01) en cultivos primarios de HAE a las 24 y 48 h después de la infección ( Fig. 3c ). La infección in vivo de ratones no demostró una pérdida de peso significativa, pero mostró una replicación viral reducida en los pulmones de la infección SHC014-CoV de longitud completa, en comparación con el SARS-CoV Urbani ( Fig. 3d, e) Juntos, los resultados establecen la viabilidad del SHC014-CoV de longitud completa, pero sugieren que se requiere una mayor adaptación para que su replicación sea equivalente a la del SARS-CoV epidémico en células respiratorias humanas y en ratones.
( a ) Esquema del clon molecular SHC014-CoV, que se sintetizó como seis ADNc contiguos (designados SHC014A, SHC014B, SHC014C, SHC014D, SHC014E y SHC014F) flanqueados por sitios únicos de BglI que permitieron el ensamblaje directo de la expresión de ADNc de longitud completa marcos de lectura abiertos (para 1a, 1b, espiga, 3, envoltura, matriz, 6–8 y nucleocápside). Los nucleótidos subrayados representan las secuencias sobresalientes formadas después de la escisión de la enzima de restricción. ( b , c ) Replicación viral de SARS-CoV Urbani (negro) o SHC014-CoV (verde) después de la infección de células Vero ( b ) o cultivos de células HAE de interfaz aire-líquido primario bien diferenciados ( c) a un MOI de 0.01. Se recogieron muestras en puntos de tiempo individuales con repeticiones biológicas ( n = 3) para cada grupo. Los datos representan un experimento para las células Vero y HAE. ( d , e ) Pérdida de peso ( n = 3 para SARS-CoV MA15, n = 7 para SHC014-CoV; n = 6 para SARS-Urbani) ( d ) y replicación viral en los pulmones ( n = 3 para SARS-Urbani y SHC014-CoV) ( e ) de ratones BALB / c de 10 semanas de edad infectados con 1 × 10 5 pfu de SARS-CoV MA15 (gris), SHC014-CoV (verde) o SARS-CoV Urbani (negro) a través del en ruta o en camino. Cada punto de datos representa la media del grupo, y las barras de error definen la sem **P <0.01 y *** P <0.001 usando la prueba t de Student de dos colas de puntos de tiempo individuales.
Durante la epidemia de SARS-CoV, se establecieron rápidamente vínculos entre las civetas de palma y las cepas de CoV que se detectaron en humanos 4 . Sobre la base de este hallazgo, el paradigma de emergencia común sostiene que la epidemia de SARS-CoV se originó como un virus del murciélago, saltó a las civetas y cambios incorporados dentro del dominio de unión al receptor (RBD) para mejorar la unión a civeta Ace2 (ref. 18 ). La posterior exposición a personas en los mercados de animales vivos permitió la infección humana con la cepa de civeta, que, a su vez, se adaptó para convertirse en la cepa epidémica ( Fig. 4a ). Sin embargo, el análisis filogenético sugiere que las primeras cepas humanas de SARS parecen estar más estrechamente relacionadas con las cepas de murciélago que con las cepas de civeta 18. Por lo tanto, un segundo paradigma argumenta que la transmisión directa murciélago-humano inició la aparición de SARS-CoV y que las civetas de palma sirvieron como huésped secundario y reservorio para la infección continua ( Fig. 4b ) 19 . Para ambos paradigmas, la adaptación de la espiga en un huésped secundario se considera una necesidad, y se espera que la mayoría de las mutaciones ocurran dentro de la RBD, lo que facilita una infección mejorada. Ambas teorías implican que los grupos de CoV de murciélagos son limitados y que las mutaciones en el rango del huésped son aleatorias y raras, lo que reduce la probabilidad de futuros eventos de emergencia en humanos.
Las cepas de coronavirus se mantienen en grupos de cuasiespecies que circulan en poblaciones de murciélagos. ( a , b ) Las teorías tradicionales de aparición de SARS-CoV postulan que los mutantes del rango del huésped (círculo rojo) representan ocurrencias aleatorias y raras que permiten la infección de huéspedes alternativos. El paradigma del huésped secundario ( a ) argumenta que un huésped no humano está infectado por un virus progenitor murciélago y, a través de la adaptación, facilita la transmisión a los humanos; La replicación posterior en humanos conduce a la cepa viral epidémica. El paradigma directo ( b) sugiere que la transmisión se produce entre murciélagos y humanos sin el requisito de un huésped intermedio; La selección se produce en la población humana con virus estrechamente relacionados que se replican en un huésped secundario, lo que permite la persistencia viral continua y la adaptación en ambos. ( c ) Los datos de virus similares al SARS quiméricos sostienen que los grupos de cuasiespecies mantienen múltiples virus capaces de infectar células humanas sin la necesidad de mutaciones (círculos rojos). Aunque pueden ser necesarias adaptaciones en huéspedes secundarios o humanos para la aparición de epidemias, si los virus que contienen espigas SHC014 se recombinan con esqueletos de CoV virulentos (círculos con contornos verdes), la enfermedad epidémica puede ser el resultado en humanos. Elementos de soporte de datos existentes de los tres paradigmas.
Aunque nuestro estudio no invalida las otras rutas de emergencia, sí argumenta a favor de un tercer paradigma en el que los grupos de CoV de murciélagos circulantes mantienen proteínas de pico « equilibradas » que son capaces de infectar a los humanos sin mutación o adaptación ( Fig. 4c ). Esta hipótesis se ilustra por la capacidad de un virus quimérico que contiene el pico SHC014 en una columna vertebral del SARS-CoV para causar una infección robusta tanto en cultivos de vías aéreas humanas como en ratones sin adaptación de RBD.
Junto con la observación de esqueletos de CoV patógenos previamente identificados 3 , 20 , nuestros resultados sugieren que los materiales de partida requeridos para las cepas emergentes similares al SARS están circulando actualmente en reservorios animales. Notablemente, aunque SHC014-CoV de longitud completa probablemente requiera una adaptación de la columna vertebral adicional para mediar en la enfermedad humana, los eventos de recombinación de alta frecuencia documentados en familias de CoV subrayan la posibilidad de una futura emergencia y la necesidad de una mayor preparación.
Hasta la fecha, las pantallas genómicas de las poblaciones animales se han utilizado principalmente para identificar nuevos virus en entornos de brotes 21 . El enfoque aquí amplía estos conjuntos de datos para examinar cuestiones de emergencia viral y eficacia terapéutica. Consideramos que los virus con el pico SHC014 son una amenaza potencial debido a su capacidad de replicarse en cultivos primarios de vías aéreas humanas, el mejor modelo disponible para la enfermedad humana. Además, la patogénesis observada en ratones indica una capacidad de los virus que contienen SHC014 para causar enfermedades en modelos de mamíferos, sin adaptación de RBD.
En particular, el tropismo diferencial en el pulmón en comparación con el SARS-MA15 y la atenuación de SHC014-CoV de longitud completa en cultivos de HAE en relación con el SARS-CoV Urbani sugieren que hay factores más allá de la unión a ACE2, incluida la procesividad de pico, la biodisponibilidad del receptor o el antagonismo de las respuestas inmunes del huésped, pueden contribuir a la emergencia. Sin embargo, se requieren más pruebas en primates no humanos para traducir estos hallazgos en el potencial patogénico en humanos.
Es importante destacar que el fracaso de la terapéutica disponible define una necesidad crítica de estudio adicional y para el desarrollo de tratamientos. Con este conocimiento, se pueden producir programas de vigilancia, reactivos de diagnóstico y tratamientos efectivos que protejan contra la aparición de CoV específicos del grupo 2b, como SHC014, y estos se pueden aplicar a otras ramas de CoV que mantienen grupos heterogéneos similares.
Además de ofrecer preparación contra futuros virus emergentes, este enfoque debe considerarse en el contexto de la pausa ordenada por el gobierno de EE. UU. Sobre los estudios de ganancia de función (GOF) 22 .
Sobre la base de modelos anteriores de emergencia ( Fig. 4a, b ), no se esperaba que la creación de virus quiméricos como SHC014-MA15 aumentara la patogenicidad. Aunque SHC014-MA15 se atenúa en relación con su SARS-CoV adaptado por ratón parental, estudios similares que examinan la patogenicidad de CoV con el pico Urbani de tipo salvaje dentro del esqueleto MA15 no mostraron pérdida de peso en ratones y una replicación viral reducida 23 . Por lo tanto, en relación con el pico Urbani-MA15 CoV, SHC014-MA15 muestra una ganancia en la patogénesis ( Fig. 1 ).
Sobre la base de estos hallazgos, los paneles de revisión científica pueden considerar que estudios similares que construyen virus quiméricos basados en cepas circulantes son demasiado arriesgados para perseguirlos, ya que no se puede excluir una mayor patogenicidad en modelos de mamíferos.
Junto con las restricciones sobre las cepas adaptadas al ratón y el desarrollo de anticuerpos monoclonales que usan mutantes de escape, la investigación sobre la aparición de CoV y la eficacia terapéutica pueden verse severamente limitadas en el futuro. Juntos, estos datos y restricciones representan una encrucijada de preocupaciones de investigación del GOF; El potencial para prepararse y mitigar futuros brotes debe sopesarse contra el riesgo de crear patógenos más peligrosos. Al desarrollar políticas en el futuro, es importante tener en cuenta el valor de los datos generados por estos estudios y si este tipo de estudios sobre virus quiméricos justifican una mayor investigación frente a los riesgos inherentes involucrados.
En general, nuestro enfoque ha utilizado datos de metagenómica para identificar una amenaza potencial que representa el CoV SHC014 similar al SARS en circulación. Debido a la capacidad de los virus quiméricos SHC014 de replicarse en cultivos de vías aéreas humanas, causar patogénesis in vivo y escapar de la terapéutica actual, existe la necesidad de vigilancia y terapéutica mejorada contra los virus circulantes similares al SARS. Nuestro enfoque también desbloquea el uso de datos de metagenómica para predecir la aparición de virus y aplicar este conocimiento en la preparación para tratar futuras infecciones por virus emergentes.
Métodos
Virus, células, infección in vitro y ensayos en placa.
El SARS-CoV (Urbani) de tipo salvaje, el SARS-CoV adaptado a ratón (MA15) y los CoV quiméricos similares al SARS se cultivaron en células Vero E6 (obtenidas del Instituto de Investigación Médica de Enfermedades Infecciosas del Ejército de los Estados Unidos), cultivadas en Eagle modificado por Dulbecco medio (DMEM) (Gibco, CA) y suero de clon fetal al 5% (FCS) (Hyclone, South Logan, UT) junto con antibiótico / antimicótico (Gibco, Carlsbad, CA). Las células DBT (laboratorio de Baric, fuente desconocida) que expresan los ortólogos de ACE2 se han descrito previamente para humanos y civetas; La secuencia de murciélago Ace2 se basó en la de Rhinolophus leschenaulti , y las células DBT que expresan murciélago Ace2 se establecieron como se describe anteriormente 8 .
Los experimentos de pseudotipado fueron similares a los que usaron un pseudovirus basado en VIH, preparado como se describió anteriormente 10 , y examinado en células HeLa (Instituto de Virología de Wuhan) que expresaban ortólogos ACE2 . Las células HeLa se cultivaron en medio esencial mínimo (MEM) (Gibco, CA) suplementado con 10% de FCS (Gibco, CA) como se describió anteriormente 24 .
Las curvas de crecimiento en Vero E6, DBT, Calu-3 2B4 y células epiteliales de las vías respiratorias humanas primarias se realizaron como se describió anteriormente 8 , 25 . Ninguna de las existencias de líneas celulares en funcionamiento se autenticó o probó para detectar micoplasma recientemente, aunque las existencias de semillas originales utilizadas para crear las existencias de trabajo están libres de contaminación. Los pulmones humanos para cultivos de AEH se adquirieron bajo los protocolos aprobados por la Junta de Revisión Institucional de la Universidad de Carolina del Norte en Chapel Hill. Los cultivos de HAE representan epitelio humano altamente diferenciado de las vías respiratorias que contiene células epiteliales ciliadas y no ciliadas, así como células caliciformes. Los cultivos también se cultivan en una interfaz aire-líquido durante varias semanas antes de su uso, como se describió anteriormente 26 .
Brevemente, las células se lavaron con PBS y se inocularon con el virus o se diluyeron simuladamente en PBS durante 40 minutos a 37ºC. Después de la inoculación, las células se lavaron tres veces y se añadió un medio nuevo para indicar el tiempo ‘0’. Se cosecharon tres o más réplicas biológicas en cada punto de tiempo descrito. No se usó cegamiento en ninguna colección de muestras ni se aleatorizaron las muestras. Todo el cultivo de virus se realizó en un laboratorio de nivel de bioseguridad (BSL) 3 con ventiladores redundantes en los gabinetes de bioseguridad, como se describió anteriormente por nuestro grupo 2 . Todo el personal usaba respiradores purificadores de aire eléctricos (Breathe Easy, 3M) con trajes Tyvek, delantales y botines y tenían doble guante.
Agrupación de secuencias y modelado estructural.
Las secuencias genómicas de longitud completa y las secuencias de aminoácidos de los dominios S1 del pico de CoV representativos se descargaron de Genbank o Pathosystems Resource Integration Center (PATRIC), alineadas con ClustalX y filogenéticamente comparadas mediante el uso de la estimación de máxima verosimilitud utilizando 100 bootstraps o por usando el paquete PhyML , respectivamente. El árbol se generó utilizando la máxima probabilidad con el paquete PhyML. La barra de escala representa sustituciones de nucleótidos. Solo los nodos con soporte de arranque superior al 70% están etiquetados.
El árbol muestra que los CoV se dividen en tres grupos filogenéticos distintos definidos como α-CoV, β-CoV y γ-CoV. Los grupos de subgrupos clásicos están marcados como 2a, 2b, 2c y 2d para β-CoV, y 1a y 1b para los α-CoV. Los modelos estructurales se generaron utilizando Modeller (Max Planck Institute Bioinformatics Toolkit) para generar modelos de homología para SHC014 y Rs3367 del SARS RBD en complejo con ACE2 basado en la estructura cristalina 2AJF (Protein Data Bank). Los modelos de homología se visualizaron y manipularon en MacPyMol (versión 1.3).
Construcción de virus quiméricos similares al SARS.
Tanto los virus de tipo salvaje como los quiméricos se derivaron de SARS-CoV Urbani o del correspondiente clon infeccioso (ic) SAR adaptado a ratón (SARS-CoV MA15) como se describió previamente 27 . Los plásmidos que contienen secuencias de espiga para SHC014 se extrajeron mediante digestión de restricción y se ligaron en el plásmido E y F del clon infeccioso MA15. El clon fue diseñado y comprado en Bio Basic como seis ADNc contiguos usando secuencias publicadas flanqueadas por sitios únicos de endonucleasa de restricción de clase II (BglI). Posteriormente, los plásmidos que contenían fragmentos de genoma de SARS-CoV quimérico de tipo salvaje y SHC014-CoV se amplificaron, escindieron, ligaron y purificaron.
Las reacciones de transcripción in vitro se realizaron para sintetizar ARN genómico de longitud completa, que se transfectó en células Vero E6 como se describió previamente 2 . El medio de las células transfectadas se cosechó y sirvió como stock de semillas para experimentos posteriores. Los virus quiméricos y de longitud completa se confirmaron mediante análisis de secuencia antes de su uso en estos estudios. La construcción sintética de mutantes quiméricos y SHC014-CoV de longitud completa fue aprobada por el Comité de Bioseguridad Institucional de la Universidad de Carolina del Norte y el Comité de Investigación de Preocupación de Uso Dual.
Declaración de Ética
Este estudio se realizó de acuerdo con las recomendaciones para el cuidado y uso de animales por la Oficina de Bienestar Animal de Laboratorio (OLAW), NIH. El Comité Institucional de Cuidado y Uso de Animales (IACUC) de la Universidad de Carolina del Norte en Chapel Hill (UNC, Número de permiso A-3410-01) aprobó el protocolo de estudio en animales (IACUC # 13-033) utilizado en estos estudios.
Ratones e infección in vivo
Se pidieron ratones hembra BALB / cAnNHsD de 10 semanas y 12 meses a los Laboratorios Harlan. Las infecciones de ratones se realizaron como se describió anteriormente 20 . Brevemente, los animales fueron llevados a un laboratorio BSL3 y se les permitió aclimatarse durante 1 semana antes de la infección. Para la infección y la vacunación con virus vivos atenuados, los ratones se anestesiaron con una mezcla de ketamina y xilazina y se infectaron por vía intranasal, cuando se desafiaron, con 50 μl de solución salina tamponada con fosfato (PBS) o virus diluido con tres o cuatro ratones por punto de tiempo, por grupo de infección por dosis como se describe en la figura leyendas.
Para ratones individuales, las anotaciones de infección, incluida la falta de inhalación de la dosis completa, el burbujeo de inóculo de la nariz o la infección a través de la boca pueden haber llevado a la exclusión de los datos del ratón a discreción del investigador; después de la infección, no se definen otros criterios de exclusión o inclusión preestablecidos. No se usó cegamiento en ningún experimento con animales, y los animales no fueron aleatorizados. Para la vacunación, se vacunó a ratones jóvenes y ancianos mediante inyección de almohadilla con un volumen de 20 μl de 0,2 μg de vacuna SARS-CoV doblemente inactivada con alumbre o PBS simulado; Los ratones fueron reforzados con el mismo régimen 22 días después y desafiados 21 días después. Para todos los grupos, según el protocolo, los animales fueron monitoreados diariamente para detectar signos clínicos de enfermedad (encorvados, pelaje con volantes y actividad reducida) durante la duración del experimento.
Todos los ratones que perdieron más del 20% de su peso corporal inicial fueron alimentados en el suelo y monitoreados adicionalmente varias veces al día, siempre que estuvieran por debajo del límite del 20%. Los ratones que perdieron más del 30% de su peso corporal inicial se sacrificaron inmediatamente según el protocolo. Cualquier ratón considerado moribundo o poco probable de recuperación también fue sacrificado humanamente a discreción del investigador. La eutanasia se realizó con una sobredosis de isoflurano y la muerte se confirmó por dislocación cervical. Todos los estudios con ratones se realizaron en la Universidad de Carolina del Norte (Animal Welfare Assurance # A3410-01) utilizando protocolos aprobados por el Comité Institucional de Cuidado y Uso de Animales de la UNC (IACUC).
Análisis histológico
Se extrajo el pulmón izquierdo y se sumergió en formol tamponado al 10% (Fisher) sin inflación durante 1 semana. Los tejidos fueron incrustados en parafina y las secciones de 5 μm fueron preparadas por el centro de histopatología del Centro Integral de Cáncer Lineberger de UNC. Para determinar la extensión de la tinción de antígeno, las secciones se tiñeron para el antígeno viral usando un anticuerpo policlonal contra la nucleocápside del SARS-CoV disponible en el mercado (Imgenex) y se puntuaron de manera ciega para la tinción de la vía aérea y el parénquima como se describió anteriormente 20 . Las imágenes fueron capturadas usando un microscopio Olympus BX41 con una cámara Olympus DP71.
Ensayos de neutralización de virus.
Los ensayos de titulación de neutralización por reducción de placa se realizaron con anticuerpos previamente caracterizados contra SARS-CoV, como se describió previamente 11 ,
. Brevemente, los anticuerpos neutralizantes o el suero se diluyeron en serie dos veces y se incubaron con 100 ufp de las diferentes cepas de clones infecciosos, SARS-CoV, durante 1 ha 37 ° C. El virus y los anticuerpos se agregaron luego a una placa de 6 pocillos con 5 × 10 5 células Vero E6 / pocillo con múltiples réplicas ( n ≥ 2). Después de una incubación de 1 h a 37 ° C, las células se cubrieron con 3 ml de agarosa al 0,8% en un medio. Las placas se incubaron durante 2 días a 37 ° C, se tiñeron con rojo neutro durante 3 horas y se contaron las placas. El porcentaje de reducción de placa se calculó como (1 – (no. De placas con un anticuerpo / no. De placas sin anticuerpo)) × 100.
análisis estadístico
Todos los experimentos se realizaron contrastando dos grupos experimentales (dos virus o cohortes vacunadas y no vacunadas). Por lo tanto, las diferencias significativas en el título viral y la puntuación histológica se determinaron mediante una prueba t de Student de dos colas en puntos de tiempo individuales. Los datos se distribuyeron normalmente en cada grupo que se comparó y tuvieron una variación similar.
Bioseguridad y bioseguridad
Los estudios informados se iniciaron después de que el Comité Institucional de Bioseguridad de la Universidad de Carolina del Norte aprobara el protocolo experimental (Título del proyecto: Generación de clones infecciosos de CoV similares a SARS de murciélago; ID de plan de seguridad de laboratorio: 20145741; ID de horario G: 12279).
Estos estudios se iniciaron antes de la Pausa de Financiación de Investigación Del Proceso Deliberativo del Gobierno de EE. UU. En Investigación de Ganancia de Función Seleccionada que Involucra Virus de Influenza, MERS y SARS . Este documento ha sido revisado por la agencia de financiación, los NIH. Se solicitó la continuación de estos estudios, y esto ha sido aprobado por los NIH.
SARS-CoV es un agente selecto. Todo el trabajo para estos estudios se realizó con procedimientos operativos estándar aprobados (SOP) y condiciones de seguridad para SARS-CoV, MER-CoV y otros CoV relacionados. Nuestras instalaciones institucionales CoV BSL3 han sido diseñadas para cumplir con los requisitos de seguridad recomendados en los laboratorios de Bioseguridad en Microbiológicos y Biomédicos (BMBL), el Departamento de Salud y Servicios Humanos de los EE. UU., El Servicio de Salud Pública, los Centros para el Control de Enfermedades (CDC) ) y los NIH. Los planes de seguridad de laboratorio se presentaron al Departamento de Salud y Seguridad Ambiental (EHS) de la UNC y los CDC y el Centro de Salud y Seguridad Ambiental de la UNC lo aprobó. Se requiere acceso con tarjeta electrónica para ingresar a la instalación.
Todos los trabajadores han sido capacitados por EHS para usar de forma segura respiradores purificadores de aire (PAPR), y se han establecido hábitos de trabajo apropiados en una instalación BSL3 y planes de vigilancia médica activa. Nuestras instalaciones CoV BSL3 contienen ventiladores redundantes, energía de emergencia para los ventiladores y gabinetes y congeladores de seguridad biológica, y nuestras instalaciones pueden acomodar bastidores de mouse SealSafe. Los materiales clasificados como agentes BSL3 consisten en SARS-CoV, cepas precursoras de Bat CoV, MERS-CoV y mutantes derivados de estos patógenos. Dentro de las instalaciones de BSL3, la experimentación con un virus infeccioso se realiza en un Gabinete de Bioseguridad de Clase II (BSC) certificado.
Todos los miembros del personal usan uniformes médicos, trajes y delantales Tyvek, PAPR y cubiertas de zapatos, y sus manos tienen doble guante. Los usuarios de BSL3 están sujetos a un plan de vigilancia médica supervisado por la Clínica de Salud Ocupacional para Empleados de la Universidad (UEOHC), que incluye una vacunación anual contra la influenza física y anual y la notificación obligatoria de cualquier síntoma asociado con la infección por CoV durante los períodos de trabajo en el BSL3. Todos los usuarios de BSL3 están capacitados en protocolos de gestión de exposición e informes, están preparados para la cuarentena y han sido capacitados para la entrega segura a un departamento local de gestión de enfermedades infecciosas en una situación de emergencia. Todos los posibles eventos de exposición son reportados e investigados por EHS y UEOHC, con informes presentados tanto a los CDC como a los NIH.
Códigos de acceso
Adhesiones
Banco de datos de proteínas
Cambia la historia
20 de noviembre de 2015
En la versión de este artículo publicada inicialmente en línea, los autores omitieron reconocer una fuente de financiamiento, financiamiento de USAID-EPT-PREDICT de EcoHealth Alliance, a Z.-LS. El error se ha corregido para las versiones impresas, PDF y HTML de este artículo. .
Referencias
-
Ge, XY y col. Aislamiento y caracterización de un coronavirus tipo SARS de murciélago que usa el receptor ACE2. Nature 503 , 535-538 (2013).
-
Yount, B. y col. Genética inversa con un ADNc infeccioso de longitud completa del coronavirus del síndrome respiratorio agudo severo. Proc. Natl. Acad. Sci. USA 100 , 12995–13000 (2003).
-
Becker, MM y col. El coronavirus de tipo SARS murciélago recombinante sintético es infeccioso en células cultivadas y en ratones. Proc. Natl. Acad. Sci. Estados Unidos 105 , 19944-19949 (2008).
-
Peiris, JS, Guan, Y. y Yuen, KY Síndrome respiratorio agudo severo. Nat. Medicina. 10 , S88 – S97 (2004).
-
Al-Tawfiq, JA y col. Vigilancia de virus respiratorios emergentes. Lancet Infect. Dis. 14 , 992–1000 (2014).
-
Él, B. et al. Identificación de diversos alfacoronavirus y caracterización genómica de un nuevo síndrome respiratorio agudo severo como el coronavirus de murciélagos en China. J. Virol. 88 , 7070-7082 (2014).
-
Li, F. Reconocimiento de receptores e infecciones entre especies del coronavirus del SARS. Res antivirales. 100 , 246-254 (2013).
-
Sheahan, T. y col. Mecanismos de la expansión del rango de hospedadores del coronavirus del síndrome respiratorio agudo severo zoonótico en el epitelio de la vía aérea humana. J. Virol. 82 , 2274–2285 (2008).
-
Yoshikawa, T. y col. Respuestas inmunes innatas dinámicas de las células epiteliales bronquiales humanas a la infección grave por coronavirus aguda asociada al síndrome respiratorio. PLoS ONE 5 , e8729 (2010).
-
Qiu, X. y col. Reversión de la enfermedad avanzada por el virus del Ébola en primates no humanos con ZMapp. Nature 514 , 47–53 (2014).
-
Sui, J. y col. Ampliación de la actividad de neutralización para bloquear directamente una vía de evolución dominante del SARS-coronavirus dirigida por anticuerpos. PLoS Pathog. 4 4 , e1000197 (2008).
-
Sui, J. y col. Efectos de los anticuerpos del dominio de unión al receptor de proteína anti-pico humano sobre el síndrome de respiración respiratoria agudo severo, la neutralización del coronavirus y la aptitud física. J. Virol. 88 , 13769-13780 (2014).
-
Rockx, B. y col. Escape de los efectos de neutralización de anticuerpos monoclonales humanos in vitro e in vivo de la aptitud del coronavirus del síndrome respiratorio agudo severo. J. Infect. Dis. 201 , 946–955 (2010).
-
Spruth, M. y col. Una vacuna de coronavirus SARS candidata a virus completo doblemente inactivada estimula las respuestas de anticuerpos neutralizantes y protectores. Vaccine 24 , 652–661 (2006).
-
Bolles, M. y col. Una vacuna de coronavirus con síndrome respiratorio agudo severo doblemente inactivado proporciona protección incompleta en ratones e induce una mayor respuesta pulmonar proinflamatoria eosinofílica en el momento de la exposición. J. Virol. 85 , 12201–12215 (2011).
-
Siegrist, C.-A. en Vaccines 6th edn. (eds. Plotkin, SA, Orenstein, WA y Offit, PA) 14–32 (WB Saunders, 2013).
-
Deming, D. y col. Eficacia de la vacuna en ratones senescentes desafiados con variantes de espiga zoonótica y epidemia de SARS-CoV recombinante. PLoS Med. 3 , e525 (2006).
-
Graham, RL, Donaldson, EF y Baric, RS Una década después del SARS: estrategias para controlar los coronavirus emergentes. Nat. Rev. Microbiol. 11 , 836–848 (2013).
-
Graham, RL & Baric, RS Recombinación, reservorios y la espiga modular: mecanismos de transmisión cruzada de coronavirus. J. Virol. 84 , 3134-3146 (2010).
- Agnihothram, S. y col. Un modelo de ratón para el subgrupo 2c de betacoronavirus usando una variante HKU5 de cepa de coronavirus de murciélago. MBio 5 , e00047-14 (2014).
-
Relman, DA Metagenómica, diagnóstico de enfermedades infecciosas e investigaciones de brotes: secuencia primero, ¿preguntas después? Mermelada. Medicina. Asoc. 309 , 1531-1532 (2013).
-
Kaiser, J. La moratoria sobre estudios de virología de riesgo deja el trabajo en 14 instituciones en el limbo. ScienceInsider http://news.sciencemag.org/biology/2014/11/moratorium-risky-virology-studies-leaves-work-14-institutions-limbo (2014).
-
Frieman, M. y col. Determinantes moleculares de la patogénesis y virulencia del coronavirus del síndrome respiratorio agudo severo en modelos de enfermedad humana en ratones jóvenes y ancianos. J. Virol. 86 , 884-897 (2012).
-
Ren, W. y col. Diferencia en el uso del receptor entre el coronavirus del síndrome respiratorio agudo severo (SARS) y el coronavirus similar al SARS de origen murciélago. J. Virol. 82 , 1899-1907 (2008).
-
Sims, AC y col. La liberación del bloqueo de importación nuclear de coronavirus del síndrome respiratorio agudo severo mejora la transcripción del huésped en las células pulmonares humanas. J. Virol. 87 , 3885–3902 (2013).
-
Fulcher, ML, Gabriel, S., Burns, KA, Yankaskas, JR & Randell, SH Cultivos celulares epiteliales de vías respiratorias humanos bien diferenciados. Métodos Mol. Medicina. 107 , 183–206 (2005).
-
Roberts, A. y col. Un coronavirus SARS adaptado a ratón causa enfermedad y mortalidad en ratones BALB / c. PLoS Pathog. 3 , e5.
Expresiones de gratitud
La investigación en este manuscrito fue apoyada por subvenciones del Instituto Nacional de Alergias y Enfermedades Infecciosas y el Instituto Nacional de Envejecimiento de los Institutos Nacionales de Salud de EE. UU. (NIH) bajo los premios U19AI109761 (RSB), U19AI107810 (RSB), AI085524 (WAM), F32AI102561 (VDM) y K99AG049092 (VDM), y por la Fundación Nacional de Ciencias Naturales de China otorga los premios 81290341 (Z.-LS) y 31470260 (X.-YG), y el financiamiento de USAID-EPT-PREDICT de EcoHealth Alliance (Z. -LS). Los cultivos epiteliales de las vías respiratorias humanas fueron apoyados por el Instituto Nacional de Diabetes y Enfermedades Digestivas y Renales del NIH bajo el premio NIH DK065988 (SHR). También agradecemos a MT Ferris (Departamento de Genética, Universidad de Carolina del Norte) por la revisión de los enfoques estadísticos y CT Tseng (Departamento de Microbiología e Inmunología, Universidad de Texas Medical Branch) por proporcionar células Calu-3. Los experimentos con los virus recombinantes SHC014 quiméricos de longitud completa se iniciaron y realizaron antes de la pausa de financiación de la investigación del GOF y desde entonces han sido revisados y aprobados para su estudio continuo por el NIH. El contenido es responsabilidad exclusiva de los autores y no representa necesariamente las opiniones oficiales de los NIH.
Información del autor
Afiliaciones
- Departamento de Epidemiología, Universidad de Carolina del Norte en Chapel Hill, Chapel Hill, Carolina del Norte, EE. UU.
- Vineet D Menachery
- Boyd L Yount Jr
- Kari Debbink
- Lisa E Gralinski
- Jessica A Plante
- Rachel L Graham
- Trevor Scobey
- Eric F Donaldson
- & Ralph S Baric
- Departamento de Microbiología e Inmunología, Universidad de Carolina del Norte en Chapel Hill, Chapel Hill, Carolina del Norte, EE. UU.
- Kari Debbink
- & Ralph S Baric
- Centro Nacional de Investigación Toxicológica, Administración de Alimentos y Medicamentos, Jefferson, Arkansas, EE. UU.
- Sudhakar Agnihothram
- Laboratorio clave de patógenos especiales y bioseguridad, Instituto de Virología de Wuhan, Academia China de Ciencias, Wuhan, China
- Xing-Yi Ge
- Y Zhengli-Li Shi
- Departamento de Biología Celular y Fisiología, Universidad de Carolina del Norte en Chapel Hill, Chapel Hill, Carolina del Norte, EE. UU.
- Scott H Randell
- Centro de Fibrosis Quística, Instituto Marsico Lung, Universidad de Carolina del Norte en Chapel Hill, Chapel Hill, Carolina del Norte, EE. UU.
- Scott H Randell
- Instituto de Investigación en Biomedicina, Instituto de Microbiología de Bellinzona, Zurich, Suiza.
- Antonio Lanzavecchia
- Departamento de Inmunología del Cáncer y SIDA, Instituto del Cáncer Dana-Farber, Harvard Medical School, Boston, Massachusetts, EE. UU.
- Wayne A Marasco
- Departamento de Medicina, Harvard Medical School, Boston, Massachusetts, EE. UU.
- Wayne A Marasco
Contribuciones
VDM diseñó, coordinó y realizó experimentos, completó el análisis y escribió el manuscrito. BLY diseñó el clon infeccioso y los virus quiméricos recuperados; SA completó ensayos de neutralización; LEG ayudó a realizar experimentos con ratones; TS y JAP completaron experimentos con ratones y ensayos de placa; X.-YG realizó experimentos de pseudotipado; KD generó cifras estructurales y predicciones; EFD generó análisis filogenético; RLG completó el análisis de ARN; SHR proporcionó cultivos primarios de HAE; AL y WAM proporcionaron reactivos de anticuerpos monoclonales críticos; y Z.-LS proporcionó secuencias y plásmidos de espiga SHC014. RSB diseñó experimentos y escribió el manuscrito.
Autores correspondientes
Correspondencia a Vineet D Menachery o Ralph S Baric .
Declaraciones de ética
Conflicto de intereses
Los autores declaran no tener intereses financieros en competencia.
Texto suplementario y figuras
Figuras suplementarias 1–6 y Tablas suplementarias 1–4 (PDF 4747 kb)
Cita este artículo
Menachery, V., Yount, B., Debbink, K. et al. Un grupo de coronavirus de murciélago circulante similar al SARS muestra potencial para la emergencia humana. Nat Med 21, 1508-1513 (2015). https://doi.org/10.1038/nm.3985
- Recibido
- Aceptado
- Publicado
- Fecha de asunto
Fuente: Descarga gratis MP3 de FUERA DE CONTROL en SoundCloud.com
